Taxonomy
The genus Rhabdias Stiles et Hassall, 1905
was erected without any diagnosis, just the type species, R. bufonis
(Schrank, 1788), was indicated. The emended diagnoses for the
genus were published by Travassos (1930), Yamaguti (1961) and
Baker (1978).
Pereira (1927) discovered the new species which he placed
into a new genus Acanthorhabdias Pereira, 1927.
The differentiation from Rhabdias Stiles et Hassall,
1905 was based on the differences in cephalic end morphology:
from 8 to 10 cone-shaped circumoral protuberances found in A.
acanthorhabdias Pereira, 1927 which have never been observed
in Rhabdias spp.
Travassos (1930) proposed the new genus Entomelas
Travassos, 1930 for 3 species previously belonging to Rhabdias
Stiles et Hassall, 1905: E. entomelas (Dujardin, 1845),
E. dujardini (Maupas, 1916) and E. chameleonis (Skrjabin,
1916). The differentiation of the two genera was based on the
differences in buccal capsule size, small in Rhabdias spp.
and enlarged in Entomelas spp. Taken into account were
the absolute dimensions of buccal capsule, without respect to
the body size of the worms. E. entomelas (Dujardin, 1845)
was indicated as type species for the genus. In the same paper
Travassos (1930) also gave brief diagnoses for the genera Rhabdias
Stiles et Hassall, 1905, Acanthorhabdias Pereira, 1927
and Entomelas Travassos, 1930.
Yamaguti (1943) described the species Rhabdias horigutii
Yamaguti, 1943 and proposed to place it into the subgenus Ophiorhabdias
Yamaguti, 1943. Differentiation from the nominative subgenus
Rhabdias was based on the absence of buccal capsule in
R. (Ophiorhabdias) horigutii Yamaguti, 1943 and its specificity
to snakes. However, the differentiation was not absolutely correct
since subgenus Rhabdias was regarded as including only
amphibian parasites. The species R. fuscovenosa (Railliet,
1899), R. vellardi Pereira, 1928, R. ophidia
Goodey, 1924 and R. labiata Pereira, 1928, all possessing
the buccal capsules, were not mentioned by Yamaguti (1943). Later
Sharpilo (1976) elevated the rank of the subgenus Ophiorhabdias
Yamaguti, 1943 to generic level.
Sharpilo (1964) transferred the species Hexadontophorus
ophisauri Kreis, 1940 from Strongylidae to Rhabdiasidae. The
similarities between H. ophisauri Kreis, 1940 and Entomelas
spp. were emphasised, however, the generic position of the species
was left unchanged and the differential diagnosis for the genus
Hexadonthophorus Kreis, 1940 was not emended.
Ballantyne and Pearson (1963) transferred Pneumonema tiliquae
Johnston, 1916 from Rictulariidae to Rhabdiasidae reffering to
the close morphological and biological affinities between the
species and other rhabdiasids. The genus Pneumonema
Johnston, 1916 was differentiated from all genera of Rhabdiasidae
on the base of presence of cuticular spines in P. tiliquae
Johnston, 1916.
Szczerbak and Sharpilo (1969) proposed the new genus Kurilonema
Szczerbak et Sharpilo, 1969 for the species K. markovi
Szczerbak et Sharpilo, 1969. The genus was distinguished from
Rhabdias Stiles et Hassall, 1905 by notably larger buccal
capsule, and from Entomelas Travassos, 1930 by the absence
of teeth.
Singh and Ratnamala (1975) described a new rhabdiasid species
Shortia shortii Singh (1975) which they placed into the
new genus Shortia Singh et Ratnamala, 1975. The
new genus was differentiated from Rhabdias Stiles et
Hassall, 1905 based on the presence of hypodermal glands - "areoles"
in S. shortii Singh, 1975.
Sharpilo (1976) proposed the new genus Paraentomelas
Sharpilo, 1976 for two species, P. dujardini (Maupas,
1916) and P. kazachstanica (Sharpilo et Vakker, 1972) previously
belonging to the genus Entomelas Travassos, 1930. The
author believed that Paraentomelas species differed from
Entomelas ones in possession of 3 onchia (vs
6 onchia in E. entomelas), swollen body cuticle and needle-like
tail tip.
The revision of the genus Entomelas Travassos, 1930
and related genera has been performed by Baker (1980). The author
synonymized the genera Kurilonema Szczerbak et Sharpilo,
1969, Hexadontophorus Kreis, 1940 and Paraentomelas
Sharpilo, 1976 with the genus Entomelas Travassos, 1930,
and the genera Ophiorhabdias Yamaguti, 1943 and Shortia
Singh, 1975 with the genus Rhabdias Stiles et Hassall,
1905. The species Entomelas chameleonis (Skrjabin, 1916)
Travassos, 1930 was transferred to the genus Rhabdias
Stiles et Hassall, 1905; three species: P. dujardini (Maupas,
1916), P. kazachstanica (Sharpilo et Vakker, 1972) and
H. ophisauri Kreis, 1940 were synonymized with E. entomelas
(Dujardin, 1845).
The synonymizations were based on the peculiarities of late
parasitic development of two Rhabdias species (Baker,
1979) (the data were extrapolated to Entomelas spp. development)
and author's attitude towards significance or insignificance of
some morphological characters (e. g. presence or absence of teeth
or buccal capsule, number of teeth, size of buccal capsule, etc.).
As the result, only 4 generic names were left valid within Rhabdiasidae
Railliet, 1915:
- Acanthorhabdias Pereira, 1927
- Entomelas Travassos, 1930
- Pneumonema Johnston, 1916 and
- Rhabdias Stiles et Hassall, 1905.
This system was accepted in the "CIH Keys to nematode
parasites of vertebrates, No. 9" (Anderson, Bain 1982).
Later, Hasegawa (1989) described a new rhabdiasid species with
quite an aberrant head end morphology and proposed the new genus
Neoentomelas Hasegawa, 1989 for it. The genus
was differentiated from Entomelas Travassos, 1930 (sensu
Baker 1980) by the presence of dorsal and ventral pseudolabia
and markedly enlarged head end bearing posterior lobes.
Lhermitte-Vallarino, Bain, Deharo et al., (2005) erected a
new genus Chabirenia Lhermitte-Vallarino, Bain,
Deharo et al., 2005 for one species, Chabirenia cayennensis,
found in buccal cavity of South-American teiid lizards. In addition
to abnormal localization, the new species possesses longitudinal
cuticular ridges similar to the synlophe of trichostrongylid nematodes.
Recently, Tkach et al. (2014) revised the system of Rhabdiasidae
based on molecular phylogenetic studies. The new genus Serpentirhabdias
Tkach, Kuzmin and Snyder, 2014 was erected for the species
parasitizing snakes. This group within Rhabdiasidae was shown
to be a monophyletic and well separated from other rhabdiasids.
The phylogenetic data supported the validity of Rhabdias
(including the species parasitic in amphibians and lizards), Entomelas
and Pneumonema. The phylogenetic relationships within
the Entomelas clade confirmed the validity of E. ophisauri,
E. dujardini and E. kazachstanika; the taxa
Hexadontophorus and Paraentomelas were shown
to be polyphyletic.
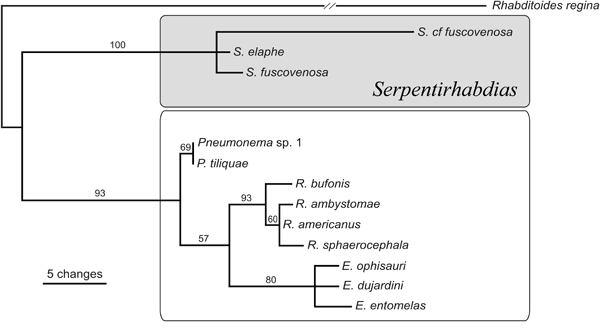
1. Phylogenetic
tree of Rhabdiasidae showing separation of Serpentirhabdias
(after Tkach et al., 2014).
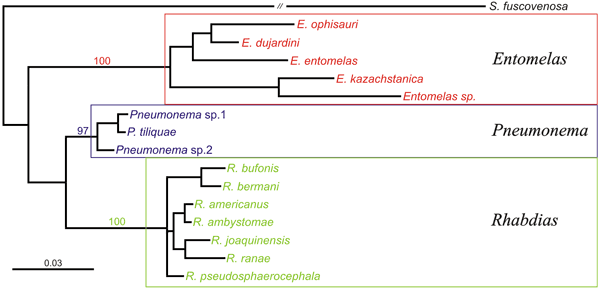
2. Phylogenetic tree of Rhabdiasidae
showing monophyly of Entomelas, Pneumonema and
Rhabdias (after Tkach et al., 2014).
Based on morphological and molecular data, the following system
of Rhabdiasidae is proposed here as the working one:
- Genus Rhabdias Stiles et Hassall, 1905
- Genus Acanthorhabdias Pereira, 1927
- Genus Chabirenia Lhermitte-Vallarino, Bain,
Deharo et al., 2005
- Genus Entomelas Travassos, 1930
- Genus Kurilonema Szczerbak et Sharpilo, 1969
- Genus Neoentomelas Hasegawa, 1989
- Genus Pneumonema Johnston, 1916
- Genus Serpentirhabdias Tkach, Kuzmin et Snyder,
2014
In our opinion, the validity of Acanthorhabdias and
Neoentomelas is to be confirmed by (molecular) phylogenetic
studies.
|