Legs in dipteran pupae are tightly
packed in a zigzag configuration. Changes in the shape or configuration
of long podomeres during eclosion have been overlooked because
they occur rapidly (in a few minutes) and the legs are hidden
inside a tight opaque confinement: the puparium in the Cyclorrhapha,
the obtect pupa in mosquitoes. We fixed insects at different times
during eclosion and obtained a temporal description of changes
in leg shape. At the start of eclosion in Calliphora vicina
and Drosophila melanogaster, femora are buckled in between
the joints. Later, the chain of podomeres straightened, pointing
posterad. Initial deformation and further stretching were passive,
exerted by forces external to the legs. The prerequisites for
this are pliability of the tubular podomeres and anchoring of
the tarsi to the confinement. Each femur was strongly crooked
instead of buckled in the mosquito Aedes cantans. The site
of bending shifted distad in the course of eclosion: a sort of
peeling. In contrast, other insects (the moth Bombyx mori,
the ants Formica polyctena and F. rufa, the honey
bee Apis mellifera) left their tight confinements without
any change in the initial zigzag leg configuration and without
transient deformations of initially straight femora and tibiae.
Larvae in holometabolous insects are adapted
for feeding and growth, legs in most of species are either short
or absent at all. Metamorphosis into the adult insect occurs
in a quiescent pupa, with immobile wing vestiges and tightly
packed legs. Pupae are often protected by silk cocoons, artificial
nest cells, or puparia (hardened larval integuments)
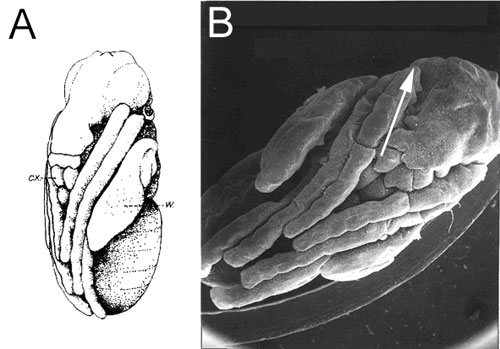
Pupa of a house fly, Musca domestica.
(A) Drawing from Hewitt (1914); (B) original SEM photograph
(specimen by L. F., SEM by J. Berger). Legs of the pupa are
packed in the zigzag configuration. The arrow indicates direction
of the hidden femur.

Z-configuration of legs in pupae. Legs
are arranged in the order: R1 (front), R2 (middle), R3 (hind),
as indicated in (F). (A) Calliphora vicina, (B) Drosophila
melanogaster, (C) Aedes cantans, head and wing vestige
ablated, tarsi recurved, (D) Bombyx mori, (E) Apis
melliphera, (F) Formica rufa. Femora and tibiae are
often bent in (A-C), but straight in (D-F), tarsi recurved in
(C).
However, flies emerge from their pupa with straighten
legs: eclosion of a mosquitoe from the film “Microcosmos”.

(click the image to watch the video in a separate
window)
There is no empty place inside the tight confinement
in order to turn the femur about 180°.

Confinements protecting the pupa: (A)
puparium in Calliphora vicina, one valve of the operculum
is shed; (B) puparium in Drosophila melanogaster, with
the pharate imago inside; (C) cocoon of Formica polyctena,
with the pupa inside; (D) hard swimming pupa of Aedes cantans;
(E) hexahedral wax cell of Apis mellifera, sealed, one
facet removed: the silk cocoon covers the cell, a small part
of the cocoon over the exposed head is removed; (F) same, the
cocoon removed, the pupa is exposed; (G) dense silk cocoon of
Bombyx mori after emergence of the moth through the orifice
at the left.

(Top row) Calliphora vicina, (middle
row) Drosophila melanogaster, (bottom row) Aedes cantans.
Leg configurations in ranked specimens demonstrate initial leg
deformations and further straightening.
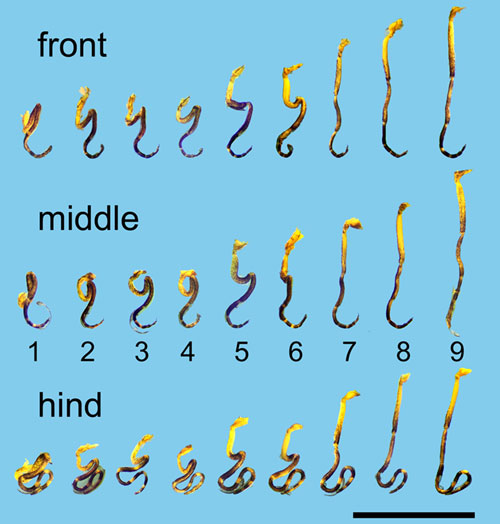
Leg configurations in eclosing imagines of
Aedes cantans. (Top row) front legs, (middle row)
middle legs, (bottom row) hind legs. Ranks are indicated in
the middle row.

Representative leg configurations in Drosophila
melanogaster, arranged by the grade of leg stretching.
(Top row) front legs, (middle row) middle legs, (bottom row)
hind legs. Ranks by the advance of the body are indicated at
the tarsi of specimens. Buckling of the hind femur occurs later
than in the front and middle legs. Legs of ten specimens of
different size are depicted. Scale bar 1 mm.
Leg ranking for Calliphora vicina is illustrated
in the separate article.
Deformations of the femur affect early ranks
and are of different type in mosquitoes and cyclorrhaphan flies.

(A) Buckling of the femur in Calliphora vicina;
(B) buckling in Drosophila melanogaster, (C) crooking
in Aedes cantans.
Position of crooking in the mosquito shifts
distally part during eclosion and thus resembles peeling.
In difference to flies, no leg deformation in
the silk worm, Bombyx mori (Lepidoptera), in the honey
bee, Apis mellifera, and in the ants Formica rufa
nd F. polyctena (Hymenoptera) occurs during their eclosion.

Partial leg stretching without podomere deformation
during eclosion in Bombyx mori. Top row – moths eclosing
from free pupae. Lower three rows – legs in the same specimens.
When the moth molts inside the cocoon, it sheds
its pupal exuvia and only afterwards dissolves the silk and
makes the opening in the cocoon: note the adult moth plus two
exuviae – the big one is the integument, the small one is the
caterpillar’s integument shed after weaving of the cocoon.

Presumably, bees and ants also left their pupal
integuments before extrication from their cocoons or wax cells.
We traced leg configurations in insects still hidden inside
their confinements.
Pupal integument in honeybees and two ant species
is soft. No traces of leg deformation or stretching of the Z-
configuration was noticed in bees. The molted bee gnaws its
way out of the wax cell.



Zigzag leg configurations in workers of Formica
polyctena, released from cocoons by adult workers.
Why for eclosion behavior and leg biomechanics
are different in Holometabola from other Arthropoda? The pupa
in holometabolans lacks any substrate for leg grasping. Presumably,
the pupal leg sheath is thin and soft at its inner side and
is torn by adult legs if the legs are hard. Or, if adult legs
are pliant, they demonstrate other types of deformation comparing
to squeezing in the rest of arthropods.


