Leonid Frantsevich, Stanislav Gorb, Vladimir Radchenko,
Dmytro Gladun, Alexey Polilov. 2014. Lehr’s fields of campaniform
sensilla in beetles (Coleoptera): functional morphology. I. General
part and allometry. Arthropod Structure & Development
43 523-535.
Abstract:
In this first of three articles
we show construction of the articular part of the elytron, the
root. The root bears a conspicuous field of campaniform sensilla
(CFS). This field was studied using light and scanning electron
microscopes. Diversity of shape of the field among beetles, types
of orientation of elongated sensilla within the field, individual
variability of their number among conspecifics are demonstrated.
Elongated CFS point to the junction of the elytron with the second
axillary plate. Presumably, they monitor twist movement in this
junction which is possible if the elytron is open. Our data on
allometric relationships between the animal size and quantitative
characteristics of the field in normally flying beetles provide
an important background for further functional analysis of this
sensory organ.
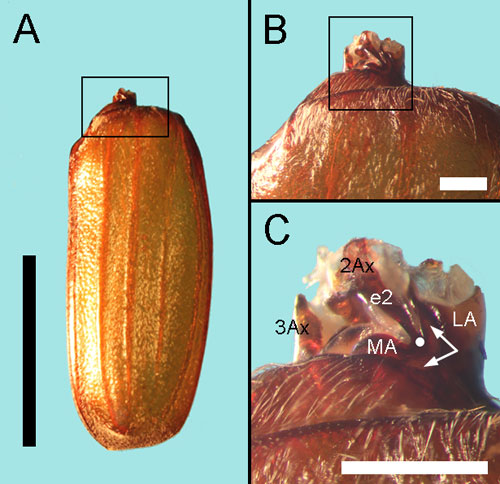
The root of the elytron in Melolontha
melolontha. (A) the whole elytron. The small process protuding
forward is the articulatory part of the elytron, i.e. the root.
The box encloses the basal part of the elytron, shown in (B).
The box in (B) encloses the root, shown in (C). The root consists
of the forked lateral apophysis (LA), the oval medial apophysis
(MA) and an intermediate sector in between where the Lehr’s field
of CFS is situated. Arrows show position of the intermediate sector.
The second and third axillary plates (2Ax, 3Ax) are embedded into
the pale articular membrane. A stripe-like elytral process (e2)
connects 2Ax with the elytron. The white dot indicates the fusion
of e2 with the intermediate sector. Scale bars 10 mm (A), 1 mm
(B, C).
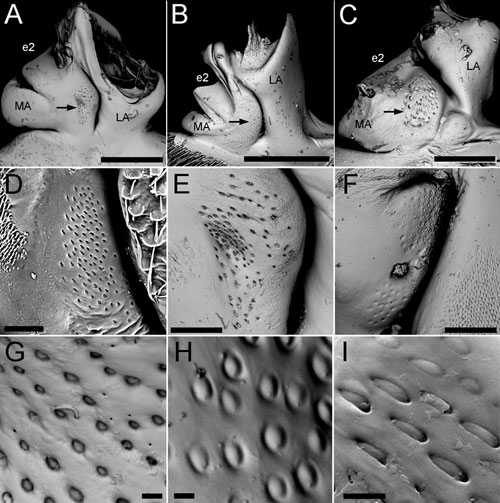
Roots and their substructures.
(A-C) general view of a root, the sensory field indicated with
an arrow; (D-F) Lehr’s fields of CFS, (G-H) campaniform sensilla.
(A, E) Hydrous piceus (Hydrophylidae); (B) Melolontha
hippocastani (Melolonthinae); (C) Lytta vesicatoria
(Meloidae); (D) Priacma serrata (Cupedidae); (F, H) Synapsis
tmolus (Scarabaeinae); (G) Cybister laterimarginalis
(Dytiscidae); (I) Ithone hexaspilota (Coccinellidae). Scale
bars 500 µm (A, B), 200 µm (C, E, F), 100 µm
(D), 10 µm (G), 5 µm (H, I).
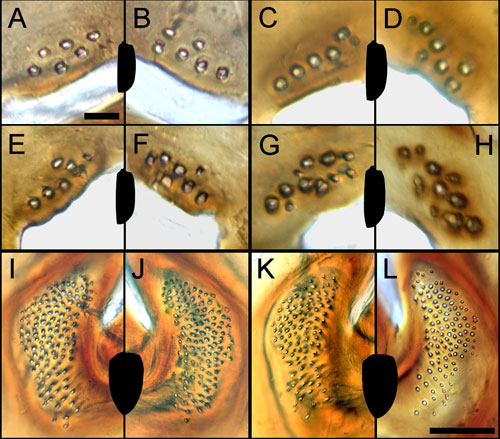
Left (A, C, E, G, I, K) and right
(B, D, F, H, J, L) fields of CFS are not exactly symmetric in
four specimens of Cicindela campestris (A-H) and in two
specimens of Anoplotrupes stercorosus (I-L). Silhouettes
of the elytra are depicted for each specimen. Pairs (G, H) and
(I, J) – females, rest are males. Scale bar for (A-H) 20 µm,
for (I-L) 100 µm. Counts of CFS for A. stercorosus
are 120 (I), 123 (J), 100 (K), and 106 (L).
We selected 14 series of flying
species belonging to the same taxon but differing in size from
big to small. The area of the sensory field is directly proportional
to the elytral area, whereas the number of sensilla is proportional
to the square root of the elytral area. Despite the great range
in the elytral area (1500 times) in series of selected species
the area of an external pit or cap of a single sensillum varies
only 25-fold. The density of sensilla per unit area of the sensory
field increases with a decrease of the elytral area.
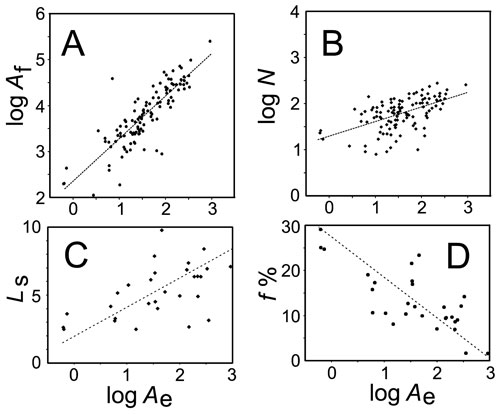
Allometric relationships versus the area of the
elytron Ae (abscissa, in mm2) for (A) the field area Af (in µ2),
(B) number N of sensilla, (C) average length Ls of the sensillum
(in µm), and (D) filling f % of the Lehr’s field with CFS
(in %). Ae, Af, and N are given in the decimal logarithmic scale.
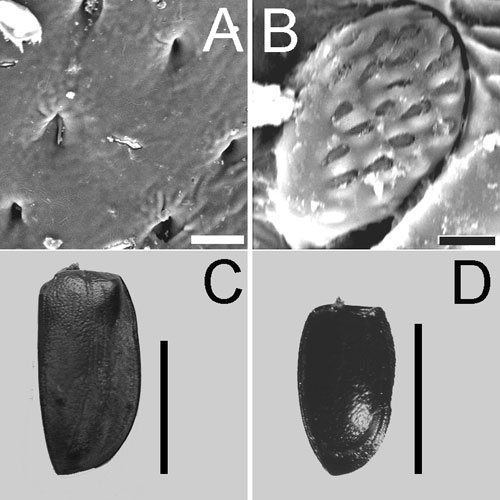
Filling of the sensory field with campaniform sensilla
(A, B) and size of the elytra (C, D) in a giant and dwarf beetle.
(A, C) Megasoma elephas (Dynastinae), (B, D) Stethorus
punctillum (Coccinellidae). Scale bars 10 µm (A), 5
µm (B), 30 mm (C), 1 mm (D). Length of the elytra differs
by 40.7 times, that of sensilla – only by 2.8 times, filling –
by 16 times.
|
|


